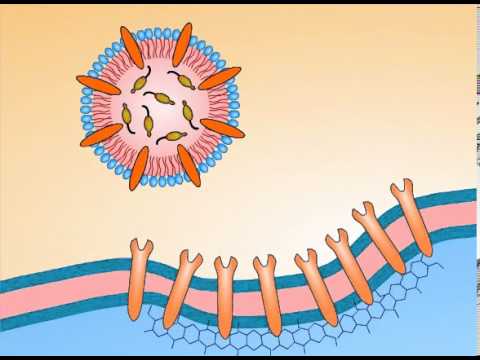
Receptor-mediated endocytosis (RME), also called clathrin-dependent endocytosis, is a process by which cells internalize molecules (endocytosis) by the inward budding of plasma membrane vesicles containing proteins with receptor sites specific to the molecules being internalized.
Process
After the binding of a ligand to plasma membrane-spanning receptors, a signal is sent through the membrane, leading to membrane coating, and formation of a membrane invagination. The receptor and its ligand are then opsonized in clathrin-coated vesicles. Once opsonized, the clathrin-coated vesicle uncoats (a pre-requisite for the vesicle to fuse with other membranes) and individual vesicles fuse to form the early endosome. Since the receptor is internalized with the ligand, the system is saturable and uptake will decline until receptors are recycled to the surface. Common for biology.
Function
The function of receptor-mediated endocytosis is diverse. It is widely used for the specific uptake of certain substances required by the cell (examples include LDL via the LDL receptor or iron via transferrin). The role of receptor-mediated endocytosis is also well recognized in the downregulation of transmembrane signal transduction. The activated receptor becomes internalised and is transported to late endosomes and lysosomes for degradation. However, receptor-mediated endocytosis is also actively implicated in transducing signals from the cell periphery to the nucleus. This became apparent when it was found that the association and formation of specific signaling complexes is required for the effective signaling of hormones (e.g. EGF). Additionally it has been proposed that the directed transport of active signaling complexes to the nucleus might be required to enable signaling as random diffusion is too slow and mechanisms permanently downregulating incoming signals are strong enough to shutdown signaling completely without additional signals transducing mechanisms.
Exocytosis is the cellular process in which intracellular vesicles in the cytoplasm fuse with the plasma membrane and release or "secrete" their contents into the extracellular space. Exocytosis can be constitutive (occurring all the time) or regulated. Constitutive exocytosis is important in transporting proteins like receptors that function in the plasma membrane. Regulated exocytosis is triggered when a cell receives a signal from the outside.
Many of the products that cells secrete function specifically for the tissue type in which the cells reside or are transmitted to more distant parts of the body. Most of these products are proteins that have gone through rigorous quality control and modification processes in the endoplasmic reticulum and Golgi membranes. It is in the trans -Golgi network, the "downstream" end of the Golgi apparatus, where cellular products are sorted and accumulate in exocytic vesicles.
Mechanisms
The mechanisms controlling regulated exocytosis were largely discovered in the 1990s. Contrary to early ideas, membranes normally do not fuse together spontaneously. This is due to the negative charges associated with the phospholipids that make up the lipid bilayer of the membranes of vesicles and organelles .
Membrane fusion requires energy and the interaction of special "adaptor" molecules present on both the vesicle and plasma membrane. The adapter molecules are highly selective and only allow vesicles to fuse with membranes of particular organelles, thus preventing harm to the cell. Once the appropriate adapter molecules bind to each other (docking), energy stored and released by ATP forms a fusion pore between the vesicle membranes and plasma membrane. The contents of the vesicle are released to the exterior of the cell (or the interior of an organelle) as the fusion pore widens. The vesicle ultimately becomes part of the plasma membrane or is recycled back to the cytoplasm.
Purpose of Exocytosis
Many cells in the body use exocytosis to release enzymes or other proteins that act in other areas of the body, or to release molecules that help cells communicate with one another. For instance, clusters of α-and β-cells in the islets of Langerhans in the pancreas secrete the hormones glucagon and insulin, respectively. These enzymes regulate glucose levels throughout the body. As the level of glucose rises in the blood, the β-cells are stimulated to produce and secrete more insulin by exocytosis. When insulin binds to liver or muscle, it stimulates uptake of glucose by those cells. Exocytosis from other cells in the pancreas also releases digestive enzymes into the gut.
Cells also communicate with each other more directly through the products that they secrete. For instance, a neuron cell relays an electrical pulse through the use of neurotransmitters . The neurotransmitters are stored in vesicles and lie next to the cytoplasmic face of the plasma membrane. When the appropriate signal is given, the vesicles holding the neurotransmitters must make contact with the plasma membrane and secrete their contents into the synaptic junction, the space between two neurons, for the other neuron to receive those neurotransmitters.
Components of the vesicle and extra neurotransmitter molecules are quickly taken up and recycled by the neuron to form new vesicles that are ready to send another pulse to an adjacent neuron. Neurons need to send many signals each second, which indicates how tight the controls are that regulate exocytosis.
The immune system also uses exocytosis to communicate information between cells. An immune cell can tell a virally infected cell that it must destroy itself to preserve other cells around it. A cell that is infected with a virus displays viral by-products on its surface, which is equivalent to the cell turning on red warning lights to attract immune cells.
Immune cells, such as the killer T cells that wander throughout the body, recognize the viral by-products and position themselves very close to the infected cell so that there is very little space between their plasma membranes. In a rapid succession, the killer T cells mobilize secretory vesicles filled with enzymes like perforin and granzyme B adjacent to the inner side of their plasma membranes. In response to a signal, the vesicles undergo exocytosis and release their contents. These enzymes then punch holes in the plasma membrane of the infected cell. This causes the cell to undergo self-destruction or apoptosis, also known as programmed cell death, to prevent further spread of the virus.
RNA interference (RNAi) technology is revolutionizing the biological discovery process as well as target discovery and validation. Using RNAi, you can turn gene expression "off", or knock it down, to better understand its function and role in disease. High-efficiency transfection is an essential first step for achieving effective gene knockdown.
Somatic cell nuclear transfer (SCNT) is a laboratory technique for creating an ovum with a donor nucleus . It can be used in embryonic stem cell research, or in regenerative medicine where it is sometimes referred to as "therapeutic cloning." It can also be used as the first step in the process of reproductive cloning.
In SCNT the nucleus, which contains the organism's DNA, of a somatic cell (a body cell other than a sperm or egg cell) is removed and the rest of the cell discarded. At the same time, the nucleus of an egg cell is removed. The nucleus of the somatic cell is then inserted into the enucleated egg cell. After being inserted into the egg, the somatic cell nucleus is reprogrammed by the host cell. The egg, now containing the nucleus of a somatic cell, is stimulated with a shock and will begin to divide. After many mitotic divisions in culture, this single cell forms a blastocyst (an early stage embryo with about 100 cells) with almost identical DNA to the original organism.
Process SCNT in stem cell research
Some researchers use SCNT in stem cell research. The aim of carrying out this procedure is to obtain stem cells that are genetically matched to the donor organism. Presently, no human stem cell lines have been derived from SCNT research.
Human Embryonic Stem cell colony on mouse embryonic fibroblast feeder layer.
A potential use of genetically-customized stem cells would be to create cell lines that have genes linked to the particular disease. For example, if a person with Parkinson's disease donated his or her somatic cells, then the stem cells resulting SCNT would have genes that contribute to Parkinson's disease. In this scenario, the disease-specific stem cell lines would be studied in order to better understand the disease.
In another scenario, genetically-customized stem cell lines would be generated for cell-based therapies to transplant to the patient. The resulting cells would be genetically identical to the somatic cell donor, thus avoiding any complications from immune system rejection.
Only a handful of the labs in the world are currently using SCNT techniques in human stem cell research. In the United States, scientists at the Harvard University Stem Cell Institute, the University of California San Francisco, and possibly Advanced Cell Technology are currently researching a technique to use somatic cell nuclear transfer to produce embryonic stem cells. In the United Kingdom, the Human Fertilisation and Embryology Authority has granted permission to research groups at the Roslin Institute and the Newcastle Centre for Life. SCNT may also be occurring in China.
In 2005, a South Korean research team led by Professor Hwang Woo-suk, published claims to have derived stem cell lines via SCNT, but supported those claims with fabricated data.Recent evidence has proved that he in fact created a stem cell line from a parthenote.
SCNT in reproductive cloning
This technique is currently the basis for cloning animals (such as the famous Dolly the sheep), and in theory could be used to clone humans. However, most researchers believe that in the foreseeable future it will not be possible to use this technique to produce a human clone that will develop to term.
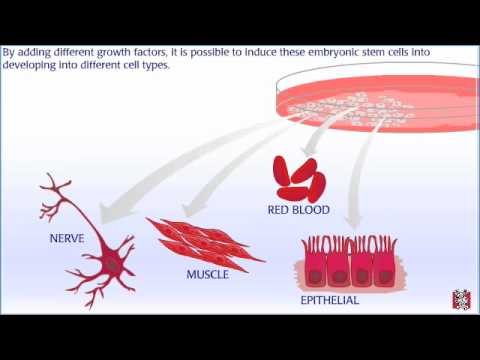
Embryonic stem cells are derived from blastocysts — embryos that are about a week old. At this stage, the blastocyst has about 100 cells. Human blastocysts like this have been donated to research from in vitro fertilization clinics. In order to get embryonic stem cell lines, scientists remove cells from the inner cell mass region. These cells have the potential to develop into any type of cell in the body. Once the cells are removed, they are placed on a culture plate with nutrients and growth factors. The blastocyst is destroyed in this process. An embryonic cell line is established when these cells multiply and divide. Under the right conditions, these cell lines can be maintained indefinitely. By adding different growth factors, it is possible to induce these embryonic stem cells into developing into different cell types. These cells could someday be used in therapies to replace damaged cells and organs.
Ligand-gated ion channels (LGICs) are a group of transmembrane ion channel proteins which open to allow ions such as Na+, K+, Ca2+, or Cl- to pass through the membrane in response to the binding of a chemical messenger (i.e. a ligand),such as a neurotransmitter.
These proteins are typically composed of at least two different domains: a transmembrane domain which includes the ion pore, and an extracellular domain which includes the ligand binding location (an allosteric binding site). This modularity has enabled a 'divide and conquer' approach to finding the structure of the proteins (crystallising each domain separately). The function of such receptors located at synapses is to convert the chemical signal of presynaptically released neurotransmitter directly and very quickly into a postsynaptic electrical signal. Many LGICs are additionally modulated by allosteric ligands, by channel blockers, ions, or the membrane potential. LGICs are classified into three superfamilies which lack evolutionary relationship: Cys-loop receptors, Ionotropic glutamate receptors and ATP-gated channels. LGICs can be contrasted with metabotropic receptors (which use second messengers), voltage-gated ion channels (which open and close depending on membrane potential), and stretch-activated ion channels (which open and close depending on mechanical deformation of the cell membrane)