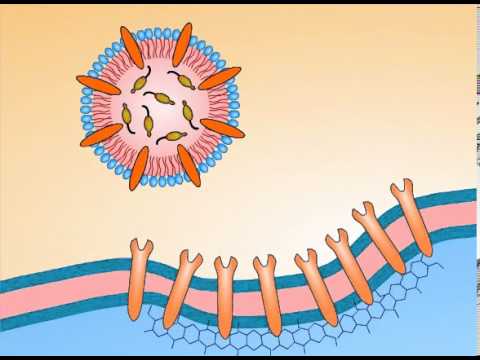
Receptor-mediated endocytosis (RME), also called clathrin-dependent endocytosis, is a process by which cells internalize molecules (endocytosis) by the inward budding of plasma membrane vesicles containing proteins with receptor sites specific to the molecules being internalized.
Process
After the binding of a ligand to plasma membrane-spanning receptors, a signal is sent through the membrane, leading to membrane coating, and formation of a membrane invagination. The receptor and its ligand are then opsonized in clathrin-coated vesicles. Once opsonized, the clathrin-coated vesicle uncoats (a pre-requisite for the vesicle to fuse with other membranes) and individual vesicles fuse to form the early endosome. Since the receptor is internalized with the ligand, the system is saturable and uptake will decline until receptors are recycled to the surface. Common for biology.
Function
The function of receptor-mediated endocytosis is diverse. It is widely used for the specific uptake of certain substances required by the cell (examples include LDL via the LDL receptor or iron via transferrin). The role of receptor-mediated endocytosis is also well recognized in the downregulation of transmembrane signal transduction. The activated receptor becomes internalised and is transported to late endosomes and lysosomes for degradation. However, receptor-mediated endocytosis is also actively implicated in transducing signals from the cell periphery to the nucleus. This became apparent when it was found that the association and formation of specific signaling complexes is required for the effective signaling of hormones (e.g. EGF). Additionally it has been proposed that the directed transport of active signaling complexes to the nucleus might be required to enable signaling as random diffusion is too slow and mechanisms permanently downregulating incoming signals are strong enough to shutdown signaling completely without additional signals transducing mechanisms.
Exocytosis is the cellular process in which intracellular vesicles in the cytoplasm fuse with the plasma membrane and release or "secrete" their contents into the extracellular space. Exocytosis can be constitutive (occurring all the time) or regulated. Constitutive exocytosis is important in transporting proteins like receptors that function in the plasma membrane. Regulated exocytosis is triggered when a cell receives a signal from the outside.
Many of the products that cells secrete function specifically for the tissue type in which the cells reside or are transmitted to more distant parts of the body. Most of these products are proteins that have gone through rigorous quality control and modification processes in the endoplasmic reticulum and Golgi membranes. It is in the trans -Golgi network, the "downstream" end of the Golgi apparatus, where cellular products are sorted and accumulate in exocytic vesicles.
Mechanisms
The mechanisms controlling regulated exocytosis were largely discovered in the 1990s. Contrary to early ideas, membranes normally do not fuse together spontaneously. This is due to the negative charges associated with the phospholipids that make up the lipid bilayer of the membranes of vesicles and organelles .
Membrane fusion requires energy and the interaction of special "adaptor" molecules present on both the vesicle and plasma membrane. The adapter molecules are highly selective and only allow vesicles to fuse with membranes of particular organelles, thus preventing harm to the cell. Once the appropriate adapter molecules bind to each other (docking), energy stored and released by ATP forms a fusion pore between the vesicle membranes and plasma membrane. The contents of the vesicle are released to the exterior of the cell (or the interior of an organelle) as the fusion pore widens. The vesicle ultimately becomes part of the plasma membrane or is recycled back to the cytoplasm.
Purpose of Exocytosis
Many cells in the body use exocytosis to release enzymes or other proteins that act in other areas of the body, or to release molecules that help cells communicate with one another. For instance, clusters of α-and β-cells in the islets of Langerhans in the pancreas secrete the hormones glucagon and insulin, respectively. These enzymes regulate glucose levels throughout the body. As the level of glucose rises in the blood, the β-cells are stimulated to produce and secrete more insulin by exocytosis. When insulin binds to liver or muscle, it stimulates uptake of glucose by those cells. Exocytosis from other cells in the pancreas also releases digestive enzymes into the gut.
Cells also communicate with each other more directly through the products that they secrete. For instance, a neuron cell relays an electrical pulse through the use of neurotransmitters . The neurotransmitters are stored in vesicles and lie next to the cytoplasmic face of the plasma membrane. When the appropriate signal is given, the vesicles holding the neurotransmitters must make contact with the plasma membrane and secrete their contents into the synaptic junction, the space between two neurons, for the other neuron to receive those neurotransmitters.
Components of the vesicle and extra neurotransmitter molecules are quickly taken up and recycled by the neuron to form new vesicles that are ready to send another pulse to an adjacent neuron. Neurons need to send many signals each second, which indicates how tight the controls are that regulate exocytosis.
The immune system also uses exocytosis to communicate information between cells. An immune cell can tell a virally infected cell that it must destroy itself to preserve other cells around it. A cell that is infected with a virus displays viral by-products on its surface, which is equivalent to the cell turning on red warning lights to attract immune cells.
Immune cells, such as the killer T cells that wander throughout the body, recognize the viral by-products and position themselves very close to the infected cell so that there is very little space between their plasma membranes. In a rapid succession, the killer T cells mobilize secretory vesicles filled with enzymes like perforin and granzyme B adjacent to the inner side of their plasma membranes. In response to a signal, the vesicles undergo exocytosis and release their contents. These enzymes then punch holes in the plasma membrane of the infected cell. This causes the cell to undergo self-destruction or apoptosis, also known as programmed cell death, to prevent further spread of the virus.
RNA interference (RNAi) technology is revolutionizing the biological discovery process as well as target discovery and validation. Using RNAi, you can turn gene expression "off", or knock it down, to better understand its function and role in disease. High-efficiency transfection is an essential first step for achieving effective gene knockdown.
Somatic cell nuclear transfer (SCNT) is a laboratory technique for creating an ovum with a donor nucleus . It can be used in embryonic stem cell research, or in regenerative medicine where it is sometimes referred to as "therapeutic cloning." It can also be used as the first step in the process of reproductive cloning.
In SCNT the nucleus, which contains the organism's DNA, of a somatic cell (a body cell other than a sperm or egg cell) is removed and the rest of the cell discarded. At the same time, the nucleus of an egg cell is removed. The nucleus of the somatic cell is then inserted into the enucleated egg cell. After being inserted into the egg, the somatic cell nucleus is reprogrammed by the host cell. The egg, now containing the nucleus of a somatic cell, is stimulated with a shock and will begin to divide. After many mitotic divisions in culture, this single cell forms a blastocyst (an early stage embryo with about 100 cells) with almost identical DNA to the original organism.
Process SCNT in stem cell research
Some researchers use SCNT in stem cell research. The aim of carrying out this procedure is to obtain stem cells that are genetically matched to the donor organism. Presently, no human stem cell lines have been derived from SCNT research.
Human Embryonic Stem cell colony on mouse embryonic fibroblast feeder layer.
A potential use of genetically-customized stem cells would be to create cell lines that have genes linked to the particular disease. For example, if a person with Parkinson's disease donated his or her somatic cells, then the stem cells resulting SCNT would have genes that contribute to Parkinson's disease. In this scenario, the disease-specific stem cell lines would be studied in order to better understand the disease.
In another scenario, genetically-customized stem cell lines would be generated for cell-based therapies to transplant to the patient. The resulting cells would be genetically identical to the somatic cell donor, thus avoiding any complications from immune system rejection.
Only a handful of the labs in the world are currently using SCNT techniques in human stem cell research. In the United States, scientists at the Harvard University Stem Cell Institute, the University of California San Francisco, and possibly Advanced Cell Technology are currently researching a technique to use somatic cell nuclear transfer to produce embryonic stem cells. In the United Kingdom, the Human Fertilisation and Embryology Authority has granted permission to research groups at the Roslin Institute and the Newcastle Centre for Life. SCNT may also be occurring in China.
In 2005, a South Korean research team led by Professor Hwang Woo-suk, published claims to have derived stem cell lines via SCNT, but supported those claims with fabricated data.Recent evidence has proved that he in fact created a stem cell line from a parthenote.
SCNT in reproductive cloning
This technique is currently the basis for cloning animals (such as the famous Dolly the sheep), and in theory could be used to clone humans. However, most researchers believe that in the foreseeable future it will not be possible to use this technique to produce a human clone that will develop to term.
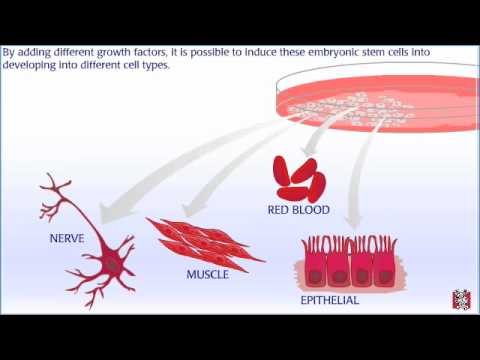
Embryonic stem cells are derived from blastocysts — embryos that are about a week old. At this stage, the blastocyst has about 100 cells. Human blastocysts like this have been donated to research from in vitro fertilization clinics. In order to get embryonic stem cell lines, scientists remove cells from the inner cell mass region. These cells have the potential to develop into any type of cell in the body. Once the cells are removed, they are placed on a culture plate with nutrients and growth factors. The blastocyst is destroyed in this process. An embryonic cell line is established when these cells multiply and divide. Under the right conditions, these cell lines can be maintained indefinitely. By adding different growth factors, it is possible to induce these embryonic stem cells into developing into different cell types. These cells could someday be used in therapies to replace damaged cells and organs.
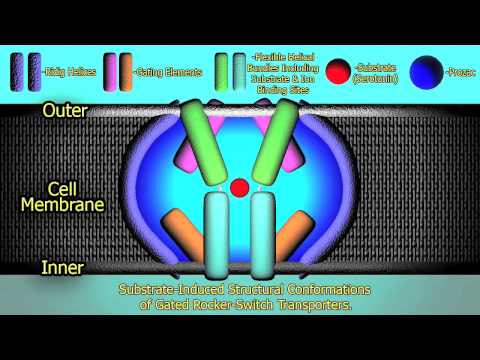
Ligand-gated ion channels (LGICs) are a group of transmembrane ion channel proteins which open to allow ions such as Na+, K+, Ca2+, or Cl- to pass through the membrane in response to the binding of a chemical messenger (i.e. a ligand),such as a neurotransmitter.
These proteins are typically composed of at least two different domains: a transmembrane domain which includes the ion pore, and an extracellular domain which includes the ligand binding location (an allosteric binding site). This modularity has enabled a 'divide and conquer' approach to finding the structure of the proteins (crystallising each domain separately). The function of such receptors located at synapses is to convert the chemical signal of presynaptically released neurotransmitter directly and very quickly into a postsynaptic electrical signal. Many LGICs are additionally modulated by allosteric ligands, by channel blockers, ions, or the membrane potential. LGICs are classified into three superfamilies which lack evolutionary relationship: Cys-loop receptors, Ionotropic glutamate receptors and ATP-gated channels. LGICs can be contrasted with metabotropic receptors (which use second messengers), voltage-gated ion channels (which open and close depending on membrane potential), and stretch-activated ion channels (which open and close depending on mechanical deformation of the cell membrane)
This animation shows how Prozac® alleviates depression. It can also be used to illustrate in general how neuron cells communicate with each other and how a neurotransmitter sends a signal from one neuron to another.
Some people with depression have a shortage of serotonin, the "mood" neurotransmitter in the brain. The antidepressant Prozac®, a Selective Serotonin Reuptake Inhibitor (SSRI), can help correct this imbalance by increasing the brain's own supply of serotonin.
This animation shows how Prozac® acts as a selective inhibitor of Serotonin Reuptake Transporter Protein, thus alleviating depression. In the brain, serotonin is associated with transmission of thoughts and feelings. In a healthy person, an optimal concentration of serotonin is available at the synapse. The imbalance of this neurotransmitter triggers emotional symptoms, like depressed mood, or physical symptoms, like aches and pains.
The blue colored layers represent the trans-membrane structure of both pre- and post-synaptic areas (the upper and lower part of the screen, respectively). Red colored masses in the post-synaptic membrane represent serotonin receptors. There are other membrane proteins as well. Depression can occur when the serotonin transporter protein (a G-protein coupled receptor; shown in white in the pre-synaptic membrane) takes up a serotonin molecule before it has a chance to bind to the post-synaptic receptor. This process is known as reuptake. Prozac® blocks the reuptake of serotonin by disabling the transporter proteins. Consequently, more serotonin molecules will be available to the post-synaptic receptor and thus depression is relieved.
A muscle contraction (also known as a muscle twitch or simply twitch) occurs when a muscle fiber generates tension through the action of actin and myosin cross-bridge cycling. While under tension, the muscle may lengthen, shorten or remain the same. Though the term 'contraction' implies a shortening or reduction, when used as a scientific term referring to the muscular system contraction refers to the generation of tension by muscle fibers with the help of motor neurons. Locomotion in most higher animals is possible only through the repeated contraction of many muscles at the correct times. Contraction is controlled by the central nervous system (CNS), which comprises the brain and spinal cord. Voluntary muscle contractions are initiated in the brain, while the spinal cord initiates involuntary reflexes.
Contractions, by muscle type
For voluntary muscles, contraction occurs as a result of conscious effort originating in the brain. The brain sends signals, in the form of action potentials, through the nervous system to the motor neuron that innervates the muscle fiber. In the case of some reflexes, the signal to contract can originate in the spinal cord through a feedback loop with the grey matter. Involuntary muscles such as the heart or smooth muscles in the gut and vascular system contract as a result of non-conscious brain activity or stimuli endogenous to the muscle itself. Other actions such as locomotion, breathing, chewing have a reflex aspect to them; the contractions can be initiated consciously or unconsciously, but are continued through unconscious reflex.
There are three general types of muscle tissues:
Skeletal muscle (voluntary and involuntary) contractions
Cardiac muscle (involuntary) contractions
Smooth muscle (involuntary) contractions.
Skeletal and cardiac muscles are called striated muscle because of their striped appearance under a microscope which is due to the highly organized alternating pattern of A band and I band.
While nerve impulse profiles are, for the most part, always the same, skeletal muscles are able to produce varying levels of contractile force. This phenomenon can be best explained by Force Summation. Force Summation describes the addition of individual twitch contractions to increase the intensity of overall muscle contraction. This can be achieved in two ways: (1) by increasing the number and size of contractile units simultaneously, called multiple fiber summation, and (2) by increasing the frequency at which action potentials are sent to muscle fibers, called frequency summation.
Multiple Fiber Summation – When a weak signal is sent by the CNS to contract a muscle, the smaller motor units, being more excitable than the larger ones, are stimulated first. As the strength of the signal increases, more motor units are excited in addition to larger ones, with the largest motor units having as much as 50 times the contractile strength as the smaller ones. As more and larger motor units are activated, the force of muscle contraction becomes progressively stronger. A concept known as the size principle allows for a gradation of muscle force during weak contraction to occur in small steps, which then become progressively larger when greater amounts of force are required.
Frequency Summation - For skeletal muscles, the force exerted by the muscle is controlled by varying the frequency at which action potentials are sent to muscle fibers. Action potentials do not arrive at muscles synchronously, and during a contraction some fraction of the fibers in the muscle will be firing at any given time. Typically when a human is exerting a muscle as hard as they are consciously able, roughly one-third of the fibers in that muscle will be firing at once, but various physiological and psychological factors (including Golgi tendon organs and Renshaw cells) can affect that. This 'low' level of contraction is a protective mechanism to prevent avulsion of the tendon - the force generated by a 100% contraction of all fibers is sufficient to damage the body.
Skeletal muscle contractions Skeletal muscles contract according to the sliding filament model:
An action potential originating in the CNS reaches an alpha motor neuron, which then transmits an action potential down its own axon.
The action potential activates voltage-dependent calcium channels on the axon, and calcium rushes in.
Calcium causes vesicles containing the neurotransmitter acetylcholine to fuse with the plasma membrane, releasing acetylcholine into the synaptic cleft between the motor neuron terminal and the motor end plate of the skeletal muscle fiber.
The acetylcholine diffuses across the synapse and binds to and activates nicotinic acetylcholine receptor on the motor end plate. Activation of the nicotinic receptor opens its intrinsic sodium/potassium channel, causing sodium to rush in and potassium to trickle out. Because the channel is more permeable to sodium, the muscle fiber membrane becomes more positively charged, triggering an action potential.
The action potential spreads through the muscle fiber's network of T-tubules, depolarizing the inner portion of the muscle fiber.
The depolarization activates L-type voltage-dependent calcium channels (dihydropyridine receptors) in the T tubule membrane, which are in close proximity to calcium-release channels (ryanodine receptors) in the adjacent sarcoplasmic reticulum.
Activated voltage-gated calcium channels physically interact with calcium-release channels to activate them, causing the sarcoplasmic reticulum to release calcium.
The calcium binds to the troponin C present on the actin-containing thin filaments of the myofibrils. The troponin then allosterically modulates the tropomyosin. Normally the tropomyosin sterically obstructs binding sites for myosin on the thin filament; once calcium binds to the troponin C and causes an allosteric change in the troponin protein, troponin T allows tropomyosin to move, unblocking the binding sites.
Myosin (which has ADP and inorganic phosphate bound to its nucleotide binding pocket and is in a ready state) binds to the newly uncovered binding sites on the thin filament (binding to the thin filament is very tightly coupled to the release of inorganic phosphate). Myosin is now bound to actin in the strong binding state. The release of ADP and inorganic phosphate are tightly coupled to the power stroke (actin acts as a cofactor in the release of inorganic phosphate, expediting the release). This will pull the Z-bands towards each other, thus shortening the sarcomere and the I-band.
ATP binds myosin, allowing it to release actin and be in the weak binding state (a lack of ATP makes this step impossible, resulting in the rigor state characteristic of rigor mortis). The myosin then hydrolyzes the ATP and uses the energy to move into the "cocked back" conformation. In general, evidence (predicted and in vivo) indicates that each skeletal muscle myosin head moves 10-12 nm each power stroke, however there is also evidence (in vitro) of variations (smaller and larger) that appear specific to the myosin isoform.
Steps 9 and 10 repeat as long as ATP is available and calcium is present on thin filament.
While the above steps are occurring, calcium is actively pumped back into the sarcoplasmic reticulum. When calcium is no longer present on the thin filament, the tropomyosin changes conformation back to its previous state so as to block the binding sites again. The myosin ceases binding to the thin filament, and the contractions cease.
The calcium ions leave the troponin molecule in order to maintain the calcium ion concentration in the sarcoplasm. The active pumping of calcium ions into the sarcoplasmic reticulum creates a deficiency in the fluid around the myofibrils. This causes the removal of calcium ions from the troponin. Thus the tropomyosin-troponin complex again covers the binding sites on the actin filaments and contraction ceases. ContractionsConcentric contraction
A concentric contraction is a type of muscle contraction in which the muscles shorten while generating force.
During a concentric contraction, a muscle is stimulated to contract according to the sliding filament mechanism. This occurs throughout the length of the muscle, generating force at the musculo-tendinous junction, causing the muscle to shorten and changing the angle of the joint. In relation to the elbow, a concentric contraction of the biceps would cause the arm to bend at the elbow and hand to move from near to the leg, to close to the shoulder (a biceps curl). A concentric contraction of the triceps would change the angle of the joint in the opposite direction, straightening the arm and moving the hand towards the leg.
Eccentric contraction
During an eccentric contraction, the muscle elongates while under tension due to an opposing force being greater than the force generated by the muscle. Rather than working to pull a joint in the direction of the muscle contraction, the muscle acts to decelerate the joint at the end of a movement or otherwise control the repositioning of a load. This can occur involuntarily (when attempting to move a weight too heavy for the muscle to lift) or voluntarily (when the muscle is 'smoothing out' a movement). Over the short-term, strength training involving both eccentric and concentric contractions appear to increase muscular strength more than training with concentric contractions alone.
During an eccentric contraction of the biceps muscle, the elbow starts the movement while bent and then straightens as the hand moves away from the shoulder. During an eccentric contraction of the triceps muscle, the elbow starts the movement straight and then bends as the hand moves towards the shoulder. Desmin, titin, and other z-line proteins are involved in eccentric contractions, but their mechanism is poorly understood in comparison to cross-bridge cycling in concentric contractions.
Muscles undergoing heavy eccentric loading suffer greater damage when overloaded (such as during muscle building or strength training exercise) as compared to concentric loading. When eccentric contractions are used in weight training they are normally called "negatives". During a concentric contraction muscle fibers slide across each other pulling the Z-lines together. During an eccentric contraction, the filaments slide past each other the opposite way, though the actual movement of the myosin heads during an eccentric contraction is not known. Exercise featuring a heavy eccentric load can actually support a greater weight (muscles are approximately 10% stronger during eccentric contractions than during concentric contractions) and also results in greater muscular damage and delayed onset muscle soreness one to two days after training. Exercise that incorporates both eccentric and concentric muscular contractions (i.e. involving a strong contraction and a controlled lowering of the weight) can produce greater gains in strength than concentric contractions alone. The caveat for this is that heavy eccentric contractions can easily lead to over-training since they are so demanding.
Eccentric contractions in movement
Eccentric contractions normally occur as a braking force in opposition to a concentric contraction to protect joints from damage. During virtually any routine movement, eccentric contractions assist in keeping motions smooth, but can also slow rapid movements such as a punch or throw. Part of training for rapid movements such as pitching during baseball involves reducing eccentric braking allowing a greater power to be developed throughout the movement.
Eccentric contractions are being researched for their ability to speed rehab of weak or injured tendons. Achilles tendinitis has been shown to benefit from high load eccentric contractions.
Isometric contraction
An isometric contraction of a muscle generates force without changing length. An example can be found in the muscles of the hand and forearm grip an object; the joints of the hand do not move but muscles generate sufficient force to prevent the object from being dropped.
Human IL-10 has been reported previously to inhibit the secretion of IFN-gamma in PBMC. In this study, we have found that human IL-10 inhibits T cell proliferation to either mitogen or anti-CD3 mAb in the presence of accessory cells. Inhibited T cell growth by IL-10 was associated with reduced production of IFN-gamma and IL-2. Studies of T cell subset inhibition by human IL-10 showed that CD4+, CD8+, CD45RA high, and CD45RA low cells are all growth inhibited to a similar degree. Dose response experiments demonstrated that IL-10 inhibits secretion of IFN-gamma more readily than T cell proliferation to mitogen. In addition, IL-2 and IL-4 added exogenously to IL-10 suppressed T cell cultures reversed completely the inhibition of T cell proliferation, but had little or no effect on inhibition of IFN-gamma production. Thus, in addition to its previously reported biologic properties, IL-10 inhibits human T cell proliferation and IL-2 production in response to mitogen. Inhibition of IFN-gamma production by IL-10 appears to be independent of the cytokine effect of IL-2 production.